This text considers the usage of legumes and rapeseed as protein source in pig nutrition. Various species of legumes are discussed, and their digestibility and nutrition contents are compared. Finally minerals are discussed, why are they needed and how to make sure inbdoors-raised pigs get all the needed minerals from their feed.
Legumes
Legumes are shrubs, herbs and trees that grow multi-leaf stalks and
reproductive flowers that produce pod-shaped fruit. The pods typically
house the pulses, which are are the seeds of a legume. Legumes are part of the pea, bean and lentil families. From legumes, only seeds are used in pig nutrition. Pulses can be used to replace soybean meal, which is the most common protein source in pig nutrition. Pulses are grinded to fit either solid or liquid feed. The most common pulses used in Europe are pea, broad bean, lupine, rape, and chickling. By-products of alcohol and bakery industries are also used as protein source.
Growing legumes diversify crop rotation, bind nitrogen and improve soil. Using pulses in pig feed usually needs no additional equipment or machinery, and can be used to increase self-sufficiency in protein feeds. However, they are sensitive to weather, and only the seeds are usable for pig feed. Most of the biomass thus goes to waste. It may also be difficult to find commercial complementary feeds for home-made pulse feed.
Nutritional content and toxic agents
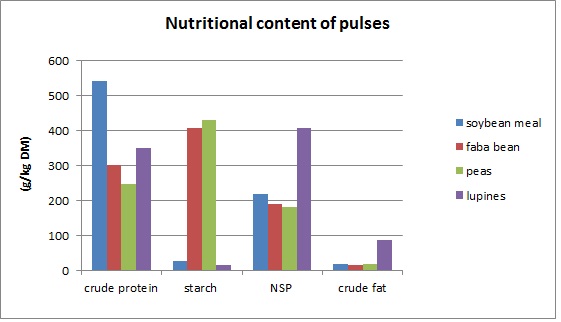
Grain legume (pea, broad bean and lupines) pulses contain mainly water-soluble protein, which has a digestibility of 85 %. They have a lot of lysine, but may lack sulphurous amino acids methionine and cystine.The higher the crude protein content in a pulse, the lower the amount of sulphurous amino acids. Compared to soy bean meal, all pulses have about 50 % less lysine and 60% less methionine and tryptophan. Intestinal digestibility of lupines equals that of soybean meal, but other pulses fall slightly behind.
Soybean meal has much as much starch as lupines. Lupines are higher in cellulose content, and may cause mild diarrhea due to their high fibre content. Faba beans and peas have seven times more starch but less sucrose than soybean.
Lupines have a much higher fat content than other pulses, soybeans included. All pulses have much polyunsaturated fatty acids, which soften the lard (pig fat) if fed in high amounts. While soft lard is healthier for humans, it makes handling and processing the carcass more difficult.
Pulses have several toxic agents.
Lectines are protease inhibitors,
inhibiting protein-degrading enzymes from working and lowering the
digestibility.They bind into the epitelial cells of the small intestine, damaging the mucuous membrane and affecting the immune system. Peas and faba beans have much less lectines than soybean meal. Other
protease-inhibitors affect the pancreatic enzymes trypsine and chymotrypsine, which split amino acids from proteins. Soybeans are high in protease-inhibitors. Heat-processing the pulses destroys the inhibitors.
Tannines may bind proteins to themselves, which inreases endogenous excretion and decreases protein digestibility. Tannines also negatively affect the taste of the feed. White-flowered legumes have no tannines. Pulses from legumes with colored flowers should be peeled, since tannines are located in the husk of the seed.
Alkaloids are toxic amines found only in lupines. They disrupt the central nervous system, decrease digestibility of all nutrients, decrease fertility and cause the feed to taste bitter. No feed should have more than 0,2-0,3 grams of alkaloids per kilogram of dry matter.
ODAP can be found only in chickling fetch/grass pea (
Lathyrys sativus). It is a neurotoxine, which in high amounts causes permanent paralysis. ODAP content can be lowered by soaking, fermenting and heat-processing the seeds. Overall,
contaminants are not a concern when using white-flowered legumes.
Less serious contaminants are vicine, saponines and alfa-galactosides. Vicine and covicine are typical for faba beans, and may affect fertility in sows. Saponines are found in legumes with colored flowes, and they taste bitter but cause no severe problems. Alfa-galactosides are carbohydrates, which pigs cannot digest at all. They are used by microbes in the colon, increasing gas production.
Using pulses in pig nutrition
Due to variable nutritional content and many contaminants, the use of pulses in pig feed must be limited. The food for sows should have a maximum of 10 % of any pulse. Piglets under 25 kg can be fed 5-15 % of pulses. Finishing pigs over 50 kg can have up to 40 % of pea, 20 % faba bean and 15 % of blue lupine in their daily feed.
Rapeseed
There are two subspecies of rapeseed: Turnip rape (
Brassica rapa) and rape (
Brassica napus). They are usually not separated in processing, and thus rape products may include turnip rape as well. Here both are referred to as rapeseed. Rapeseed is used in pig nutrition as compressed cakes, powdered seeds or groats. Organic production relies heavily on rapeseed for protein, and they also include a lot of necessary phosphorus and healthy fatty acids. Organic producers cannot however use rapeseed meals which are produced using ether extraction. Rapeseeds are high in fibre content, so they fit especially well for sows.
Rapeseeds contain two toxic agents: glucosinolates and erucic acid. Only the so-called 00-variants of rape are free from both compounds.
Glucosinolates are sulphurous, aliphatic compounds found in cruciferous plants. Glucosinolates cause bitter taste, decrease the metabolism of iodine in the thryroid gland and may damage the liver if their amount in feed exceeds 7 μmol/g. Crops cultivated in cool and humid environment develop less toxic agents
than crops in hot and dry environments. When rapeseeds are processed,
most of the glucosinolates stay in the pressed cakes.They can also be partially destroyed by heat-processing. The effects of erucic acid are controversial and not very well known.
Soybean meals can be entirely replaced with rapeseed groats or heat-processed, pressed rapeseed cakes for growing meat pigs. Only a third of soy can be replaced with rapeseed for sows. Rapeseed-fed sows lose less weight after parturition and their piglets weigh more when born and when weaned.
Minerals
From minerals, calcium and phosphorus are the most important for pigs since they are needed for muscle contractions, nervous system, energy metabolism and blood clotting. Strong bones require the right ratio of Ca and P. Phosphorus is absorbed from the small intestine with the help of Na-transferrers, which again need vitamin D to work. Absorption of Calcium aso needs vitamin D. If the animal gets more phosphorus than it needs or the phosphorus is in an insoluble form, the excess will be secreted in urine.
Lack of Ca, P and vitamin D causes rickets in young animals and osteomalacia in old. Pigs need to get 1,2-1,4 times more Ca than P. Exceeding the needed amount of Ca decreases the absorbancy of zinc and can cause skin problems for young pigs. Pregnant sows with calcium deficiency have weak contractions, and piglets are born with oxygen-deficieny and covered in feces. The probability of uterus infection is also increased.
50-90 % of the phosphorus in plants exists as
phytic acid (inositol hexakisphosphate). Compressed rapeseed and rapeseed groats contain most free
phosphorus, approximately 11 g P / kg dry matter (DM), but as much as 32
grams of phytic acid in a kilogram of DM. Barley, wheat and oats
contain only 3,5 g P / kg DM and 10 g phytic acid / kg DM. Phytic acid exists as K- and Mg-salts, and forms complexes with other positively charged ions. Monogastric animals like swine cannot digest phytic acid. They need an enzyme called
phytase to remove phosphate from the inositol ring in the phytate molecule. Often phytase is added to pig feed, because the natural phytase in plants is destroyed in heat-processing. Some by-products of alcohol industry (barley protein feed) have almost only free P, since the phytic acid denaturates in the ethanol creation process.
Grains
Grains,
and especially barley, is the most important component in pig nutrition
in Europe. Barley can be given to pigs of all ages, and without mixing
it to other ingredients. Oats are very fibrous, and are used mostly for
sows to replace 50% of the barley. For meat pigs, the diet must contain
less than 50% of wheat and oats to avoid softening the lard. Rye is not
commonly used at all, but may be used in small proportions for growing
pigs.
All grains must be fed either dried or as silage, but the grains must be
flattened, grinded or powdered before feeding. Grinded grains preserved
with propionic acid are safe to feed, since the acid is a natural
product of the gastric system, and it keeps the grain free from spores,
fungi and mold. Anaerobic preservation can also be used. Grains must
always be clean and in good condition before feeding. Moldy grains cause
gastric problems or other severe effects, and sprouted and very light
grains have only low nutritional value. Moldy grains cannot be given to
sows and piglets at all.
All grains contain roughly the
same amount of starch, ash and crude protein. Whole-grain oats and
barley have the most crude carbohydrates (hemicellulose, cellulose and
lignin), but peeled oats and barley have less CC than whole-grain wheat
and rye. Compared to soybean meals, grains contains approximately 2/3
less protein. The amino acids in grains have intestinal digestibility of
60-90 depending on the amino acid. Grains fertilized with nitrogen
contain 2 % more protein, but less lysine than non-nitrogen-fertilized
grains.

Grains
must be harvested at the right time and silaged and grinded properly.
Good grains are clean, bright-colored and smell fresh. A hectolitre of
barley should weight over 66 kgs and oat over 56 kgs. Lighter grains
have less organic matter and considerably more neutral detergent fibres,
which make the grains less digestible. All grains must be dried to 14 %
of moisture immediately after harvesting to ensure microbiological
quality. The protein content must be analyzed and found to be around 12-13 %. Barley must contain over 59 % of starch (in dry matter). Grains should be stored so that birds, pigs or pests cannot contaminate them with feces.
More information
Plants poisonous to livestock (Cornell University):
http://www.ansci.cornell.edu/plants/php/plants.php?action=display&ispecies=swine
Jezierny, D.; Mosenthin, R.; Bauer, E. 2010. The use of grain legumes as a protein source in pig nutrition: A review.
Animal Feed Science and Technology
vol. 157 issue 3-4 May 11, 2010. p. 111-128
Partanen K., Valaja J., Jalava T., Siljander-Rasi H. 2001.
Composition, ileal amino acid digestibility and nutritive value of organically grown legume seeds and conventional rapeseed cakes for pigs. Agricultural and food science in Finland, Vol 10 (2001): 309-322.







%20copy.JPG)
.jpg)














{kind=link}
{kind=link}
{kind=link}